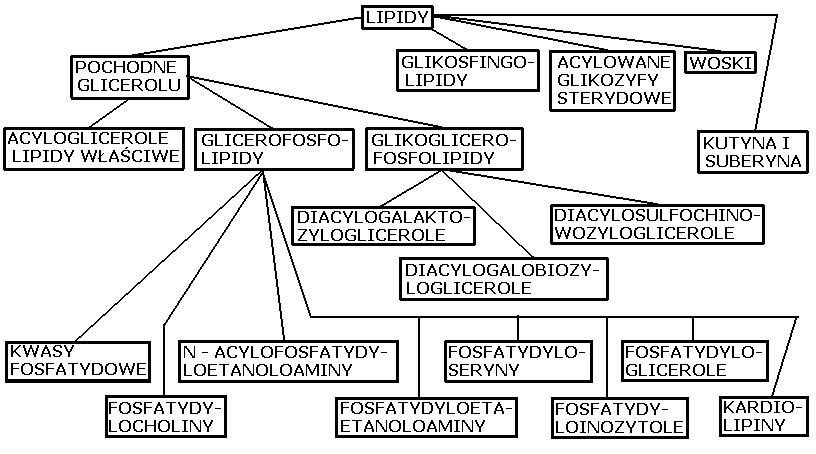
LIPIDY
![]()
fosfolipidy
- glikolipidy - sterole
Lipidy to liczna grupa związków organicznych o różnorodnym składzie i budowie, których cząsteczki zawsze zawierają długołańcuchowe kwasy tłuszczowe, co nadaje im wspólną właściwość – złą rozpuszczalność w wodzie, natomiast dobrą w rozpuszczalnikach organicznych. Jest to związane z występowaniem w ich strukturze dużej liczby hydrofobowych grup węglowodorowych (-CH2-), tworzących długie alifatyczne łańcuchy, które pochodzą od kwasów tłuszczowych, aldehydów, alkoholi, pochodnych sfinganiny lub cząsteczek steroli. Oprócz hydrofobowych łańcuchów węglowodorowych cząsteczki lipidów zawierają w swoim składzie hydrofilowe grupy polarne np.: estrowe, fosforanowe, wodorotlenowe, aminowe i inne. Grupy te zdolne są do oddziaływania z wodą przez tworzenie wiązań wodorowych. Związki, które zawierają w swoim składzie zarówno część hydrofilową jak i hodrofobową nazywa się związkami amfifilowymi. Właściwości amififilowe zapewniają lipidom, a w szczególności fosfolipidom, dużą aktywność powierzchniową. Lipidy adsorbują się na powierzchni wody zmniejszając jej napięcie powierzchniowe.
Rozróżnia się lipidy prosta i lipidy złożone. Do lipidów prostych należą triglicerydy – estry glicerolu z kwasami tłuszczowymi, stanowiące głównie materiały zapasowe (surowce energetyczne) organizmu, a u zwierząt także podściółkę i warstwę ochronną różnych narządów,
woski naturalne – estry alkoholi z kwasami tłuszczowymi, tworzące warstwy chroniące przed nadmiernym parowaniem wody, a także estry sterolowe (kwasów tłuszczowych z cholesterolem), do lipidów prostych są często zaliczane wyższe kwasy tłuszczowe (których łańcuch zawiera 10 atomów węgla lub więcej), alkany, alkanole, karotenoidy i sterole; związki te są zaliczane do lipidów ze względu na podobne zachowanie się w procesie przemiany materii oraz podobną rozpuszczalność. Lipidy złożone zawierają w cząsteczce oprócz kwasów tłuszczowych i alkoholi (glicerolu lub sfingozyny) kwas fosforowy, cholinę, etanoloaminę, sacharydy (np.galaktozę) i inne. Należą do nich tak zwane lipidy narządowe: fosfolipidy i glikolipidy, wchodzące w skład elementów strukturalnych komórek (błon i ziarnistości), a u zwierząt występujące w szczególnie dużej ilości w tkance mózgowej i włóknach nerwowych. W organizmie lipidy występują przeważnie w postaci połączeń z białkami (lipoproteiny).
Fosfoglicerydy
są fosfolipidami, których budowa oparta jest na glicerolu (trójwęglowym
alkoholu) stanowiącym rdzeń strukturalny cząsteczki fosfoglicerydu. Budowę
strukturalną cząsteczek najważniejszych fosfoglicerydów (fosfatydylocholiny
, fosfatydyloetanoloaminy, fosfatydyloseryny, fosfatydyloinozytolu). Grupy
hydroksylowe glicerolu tworzą wiązania estrowe z grupami karboksylowymi kwasów
tłuszczowych i kwasu fosforowego.
Sfingolipidy
są fosfolipidami, których budowa oparta jest na sfingozynie - długołańcuchowym
nienasyconym alkoholu. Cząsteczka sfingozyny, z dołączonym do grupy aminowej
kwasem tłuszczowym, nosi nazwę ceramidu. Najczęściej występującym w błonach
przedstawicielem sfingolipidów jest sfingomielina , która występuje głównie
w tkance nerwowej. Za pomocą związków znaczonych izotopami fosforu
stwierdzono, że przemiany sfingomieliny są tam bardzo powolne. Jest ona też
mniej niż lecytyny i kefaliny podatna na działanie czynników chemicznych.
Okazuje się, że zawiera bardzo mało nienasyconych kwasów tłuszczowych, związanych
peptydowo (poprzez azot) z cząsteczką sfingozyny. Te właściwości
sfingomieliny świadczą o tym, że pełni ona funkcje izolatora tkanki
nerwowej. Występuje także w tkankach nerek, krwi, i w mielinie.
Fosfolipidy mają kluczowe znaczenie w fizjologii i
patologii błon biologicznych. Stanowią one podstawowy strukturalny i czynnościowy
budulec wszystkich błon komórkowych i struktur błonowych organelli wewnątrzkomórkowych.
Wpływają korzystnie na elastyczność, płynność i przepuszczalność błon
komórkowych. Utrzymują i przywracają odpowiednią aktywność receptorów i
białek związanych z błonami. Mają decydujące znaczenie dla utrzymania
prawidłowej aktywności biologicznej układów enzymatycznych związanych z błonami,
takich jak np. cyklaza adenylowa, lipaza lipoproteinowa, ATP- aza sodowo -
potasowa, ATP - aza wapniowa, acylotransferaza lecytyna: cholesterol, oksydaza
kwasu bursztynowego, transferaza glukozylowa czy fosforylaza. Biorą udział w
utrzymaniu i regulacji procesów transportu czynnego i biernego przez błony.
Odgrywają istotną rolę w powstawaniu potencjałów błonowych. Regulują zależne
od błon procesy metaboliczne, które zachodzą pomiędzy przestrzenią wewnątrz-
i zewnątrzkomórkową. Są niezbędne w procesach różnicowania, proliferacji
i regeneracji komórek. Są substratem do produkcji wielu ważnych związków
chemicznych, ponieważ zawierają w swoim składzie wielonienasycone kwasy tłuszczowe,
cholinę i resztę kwasu fosforowego. Wielonienasycone kwasy tłuszczowe są ważnymi
prekursorami cytoprotekcyjnych prostaglandyn, leukotrienów i innych eikozanoidów,
a reszta kwasu fosforowego odgrywa duże znaczenie m.in. w produkcji ATP. Są
prekursorami tzw. drugich przekaźników uwalnianych przez fosfolipazę C -
diacyloglicerolu i trifosforanu inozytolu, biorących udział w wydzielaniu
wielu hormonów (m.in. katecholamin, insuliny, parathormonu, kalcytoniny) oraz
powstawaniu czynników wzrostu. Są emulgatorami tłuszczów w przewodzie
pokarmowym i przyczyniają się do lepszego ich trawienia oraz wchłaniania
witamin w nich rozpuszczalnych (A, D, E, K). Są ważnymi emulgatorami w żółci.
Mają wpływ na reakcje immunologiczne na poziomie komórkowym. Są także
strukturalną i funkcjonalną częścią krążących lipoprotein.
Wykazano, że różne czynniki (toksyczne,
metaboliczne, wirusowe, immunologiczne itp.) mogą powodować uszkodzenia błon
hepatocytów, co w rezultacie prowadzi do zaburzenia funkcji komórek lub ich śmierci.
Dowiedziono, że prawie każde uszkodzenie wątroby wiąże się z uszkodzeniem
na poziomie błon biologicznych hepatocytów. Zbyt mała ilość egzogennych
fosfolipidów w diecie zawierających w swoim składzie wielonienasycone kwasy tłuszczowe
uniemożliwia pokrycie ich zwiększonego zapotrzebowania w przypadku choroby, a
synteza wewnątrzustrojowa jest albo bardzo energochłonna (lecytyna), albo
niemożliwa (polienylofosfatydylocholina). W wielu chorobach istnieje konieczność
podawania egzogennych fosfolipidów w postaci zawierających je preparatów
farmakologicznych.

Przykładem glikolipidów są glikozylodwuglicerydy, które składają się z mono-, dwu- lub trój sacharydów połączonych wiązaniem glikozylowym z grupami hydroksylowymi dwuglicerydu. Lipidy te występują w chloroplastach (np. monogalaktozylodwuacyloglicerol), i błonach bakterii gramdodatnich (np. dwusacharylodwuacyloglicerole ). Inne glikolipidy tej grupy występują ponadto w składzie lipidów mózgu i tkanki nerwowej, śledziony, nerek, płucach, wątroby i erytrocytów.
Do grupy glikolipidów należą również gangliozydy i glikozydy sterylowe. Gangliozydy są złożonymi cerebrozydami (występującymi głównie w mózgu), w których reszta ceramidowa przyłączona jest do reszty cukrowej zawierającej galaktozaminę i kwas sialowy. Lipidy te występują najczęściej w zewnętrznych błonach komórek. Cechą charakterystyczną glikozydów sterylowych jest wiązanie glikozylowe cząsteczki monosacharydu z grupami OH sterolu. Lipidy te występują w błonach roślinnych i pełnią tam rolę materiału budulcowego i izolacyjnego. Biorą także udział w procesach dotyczących działalności układu nerwowego. Ważną funkcję we wspomganiu układu nerwowego pełnią cerebrozyty występują głównie w mózgu. W innych tkankach występują bardzo podobne związki zwane sulfatydami. Ich charakterystycznym składnikiem jest kwas siarkowy, związany estrowo z C – 6 galaktozy. Jeszcze bardziej złożoną budowę mają gangliaydy, gdyż zawierają kilka odmian powiązanych heksoz i ich pochodnych.

Sterole
